納米顆粒,自由基和氧化應激
自由基是外層有自由電子的不穩定原子或分子。這使得它們具有很高的反應性,因為自由電子總是努力形成穩定的鍵。這種穩定包括從另一個分子中獲得一個電子,從而引發連鎖反應。這種反應在人體內無所不在,但在某些情況下會破壞生物分子。納米顆粒是否被細胞內吸收導致自由基產生的激活,目前正在討論中。
正在進行的研究正在調查納米顆粒表麵形成的自由基數量是否足以誘導細胞效應。這份檔案概述了自由基是什麼,它們如何產生,為什麼生物需要它們,它們如何被中和,以及我們所知道的納米顆粒和自由基產生之間的聯係。
簡介
人們一致認為,納米顆粒的表麵結構在與細胞的相互作用中起著至關重要的作用,因此在評估其健康影響時應予以考慮。討論通常集中在納米顆粒吸收進入細胞後自由基發展的作用機製上。
自由基在氧依賴生物係統中起著重要作用。它們是細胞呼吸和其他重要細胞過程的重要組成部分,但也參與衰老和疾病發展。
自由基是外層有自由電子(未成對電子)的不穩定分子。它們是高度反應性的,因為自由電子總是努力與其他電子成鍵,形成共價對。在這個過程中,自由基從其他分子中剝奪電子。這種連鎖反應無處不在,存在於人體的每個細胞中。除了影響細胞調節外,它還會破壞碳水化合物、脂肪、蛋白質和核酸等分子。
什麼是自由基?
自由基是含有一個或多個未成對電子的原子或分子,從這個意義上說,是“自由的”。這使得許多自由基具有高度活性,即它們有很強的成對傾向,以抵消不穩定的未成對情況。
為此,自由基從任何可用的供體獲得電子,或將一個電子提供給合適的受體,後者又被修飾成次生自由基。這種連鎖反應會造成生物損傷(圖1)。
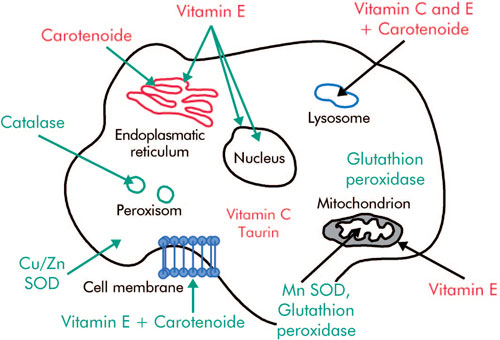
在細胞和環境中,自由基有許多不同的來源。在有氧生物中,自由基是在正常代謝過程中或通過正常代謝過程產生的。
主要來源包括質膜中的電子轉移和線粒體膜中的細胞呼吸作用。生產過程可以通過酶(用催化劑)進行,也可以通過非酶進行。
一個研究小組聲稱3.線粒體是氧化損傷的主要來源,因為自由基如超氧化物可以從電子傳遞鏈中逃逸。
大約3- 10%的氧氣沒有被完全處理,即被還原。它們可以進入細胞的細胞質,在那裏自由基可以與其他物質發生反應,從而通過從這些物質中去除電子形成新的自由基。這引發了一個連鎖反應,電子會改變它們的主人,最終導致細胞變化,如DNA修飾或酶破壞。
抗氧化劑-氧化應激
為了對抗自由基對細胞內的破壞,細胞發展出了一種所謂的細胞內抗氧化係統。這個過程通過蛋白質(酶)將自由電子轉化為非反應形式。
抗氧化劑通過抑製、延遲或阻礙物質的氧化來調節氧化反應4。細胞內的酶起著抗氧化劑的作用,是細胞防禦係統的支柱5、6.
關鍵的抗氧化酶含有某些屏蔽和保護蛋白質的元素7、8.
非酶促抗氧化劑也可以中和自由基4(例如水溶性物質如維生素C、穀胱甘肽或脂溶性物質如維生素E或維生素A/β胡蘿卜素)。
例如,SOD酶將超氧化物自由基轉化為過氧化氫,然後通過催化分解為水和氧。
自由基不僅是破壞代謝的產物,而且具有一係列重要的功能。例如,它們在免疫防禦中發揮作用,因為白細胞和巨噬細胞利用它們的殺菌作用:它們產生自由基,從而消滅細菌和其他外來物質。
此外,自由基可能通過介導細胞程序性死亡(凋亡)在機體的腫瘤抑製中發揮作用。
免疫相關細胞也利用活性氧的反應電位作為細胞防禦機製,防止進入病原體,殺死細菌、病毒和退化細胞。
自由基還履行重要的生理功能,如調節血管張力和那些受氧濃度控製的細胞功能。它們還影響信號傳遞機製,引發氧化應激反應和細胞凋亡11.
細胞防禦機製的能力是有限的。因此,氧化應激會導致功能障礙,甚至導致細胞死亡。氧化應激是細胞內產生的自由基和細胞防禦機製之間不平衡的結果。
自由基的增加或抗氧化物質的減少會破壞氧化劑和抗氧化劑之間的平衡。氧化應激會引發許多潛在的破壞性生化反應9.
研究表明,自由基的產生直接參與了油脂、蛋白質和核酸等大分子的氧化破壞。在一定條件下,吞噬體(例如,細菌被吸收的囊泡)10大量的巨噬細胞可以退化並將其內容物釋放到其他細胞室,通過氧化反應破壞DNA。
因此,慢性感染可通過促進巨噬細胞的永久性吞噬活性來引發慢性炎症反應。
此外,眾所周知,慢性炎症是誘發癌症的風險之一。
此外,自由基在許多退行性疾病和細胞老化過程中發揮作用3..
細胞內氧化還原平衡(氧化還原穩態)
自由基及其衍生物,以及可歸因於自由基的活性非自由基,總是以相對較低和平衡的數量存在於生命係統中。
自由基的濃度取決於它們的產生和清除。清除是由各種酶和非酶抗氧化劑控製的(例如維生素E, A, C和穀胱甘肽)。
當ROS產生速率和抗氧化能力達到平衡時,細胞處於穩定狀態。這被稱為平衡氧化還原容量。這種平衡可能會被ROS產量的增加或抗氧化劑能力的降低所破壞。
由於自由基可以將電子提供給合適的受體(還原反應)或將未配對的電子與合適的供體(氧化反應)結合,自由基在維持細胞內氧化還原平衡方麵發揮著重要作用。
根據不平衡的持續時間和強度(可以暫時限製),細胞的氧化還原調節實現了一種代償功能。這種生理機製被稱為氧化還原穩態。
然而,當自由基的持續產生被觸發時,例如氧化應激,氧化還原內穩態就會變得不平衡,因為細胞機製不再能夠建立正常水平。這可以持續地改變信號傳輸,但也會導致細胞的基因和蛋白質水平的變化,從而促進所謂的氧化條件或過程。
這幾乎包括了所有能獲得單個電子的複雜分子(DNA、蛋白質、脂類和碳水化合物),從而被高活性自由基破壞。當活性氧在較長時間內持續升高(慢性疾病)時,自由基會造成損傷並導致病理狀況。
納米顆粒和自由基
各種各樣的在體外而且在活的有機體內研究表明,納米粒子(富勒烯、碳納米管、量子點、發射粒子)可以觸發自由基的形成。12、13.納米顆粒可被某些細胞(巨噬細胞)主動(吞噬)並啟動ROS的形成14、15.細胞對顆粒的被動吸收也有文獻記載。
然而,決定性的問題是,當更多的粒子被吸收時,每個細胞是否會形成更多的ROS。納米顆粒傾向於形成聚集/團聚。目前尚不清楚它們在這種配置下是否能產生較高的ROS水平。
ROS也可以直接在顆粒表麵形成,盡管這取決於顆粒結構(例如,金屬顆粒起催化劑的作用)。由於小顆粒的比表麵積與質量比更大,可以形成更多的ROS比大顆粒16、17.目前尚不清楚聚集是否影響活性氧形成的數量。
活性氧的過量產生或慢性產生會引起炎症反應、組織變化和DNA、蛋白質和脂質損傷。納米顆粒還會引起細胞內的機械損傷,從而引發氧化應激。
抗氧化劑在一定程度上可以通過細胞的內穩態活性來中和自由基。如果產生的活性氧多於中和的活性氧,那麼這個係統就會發生變化,某些生物分子就會被氧化和/或改變。
在一項初步研究中18一種特殊的小鼠模型顯示,長(約20米)針狀納米管注入腹腔(腹腔內)會引起慢性炎症,而短的和/或彎曲的以及長彎曲的納米管不會引起這種效應。
由於這些管的結構(形式、長度和溶解度)與石棉纖維非常相似,因此正在討論一種類似的作用機製。石棉暴露會導致胸膜區間皮瘤(結締組織腫瘤)。
這種腫瘤不一定是惡性的。當巨噬細胞試圖消化吸收的針狀纖維時,它們就會形成。由於纖維太長,這些細胞無法生長。
因此,自由基的存在伴隨著所謂的巨細胞的形成,因為幾個細胞相互融合,以成功地吸收纖維。這些細胞的長期激活導致結節狀新組織的形成,即所謂的肉芽腫。隨著時間的推移,這些會發展成間皮瘤。
上述研究特別使用了在形狀和長度上類似石棉纖維的納米管。然而,為了比較它們的效果,也測試了其他纖維。正如預期的那樣,結果顯示,隻有長針狀的納米管,而不是短的和/或彎曲的納米管,觸發了慢性炎症(肉芽腫)。
評估這項研究的一個重要考慮因素是每隻小鼠相對較高的納米管濃度(50 μ g)和所使用的非常特定的動物模型。研究本身也強調了納米管在預處理方麵的差異。
此外,為了評估這項研究的意義,必須考慮實驗條件和使用的動物數量有限。雖然必須認真對待這項研究的信息,但它需要得到核實和複製。
結論
被細胞內吸收的納米顆粒可以誘導細胞效應,其生物學相關性仍有待澄清。
實驗研究表明,納米顆粒可以引發自由基的產生。這種活性分子的長期釋放會導致組織變性。
大多數研究都在相對較短的暴露時間內測試了非常高濃度的納米顆粒。這些數據妨礙了對健康風險進行確定的評估。細胞和有機體的內穩態活性抵消了暴露於納米顆粒的影響。
目前還不清楚該係統在什麼時候會變得不平衡,從而導致生物和健康方麵的影響。這需要對潛在機製進行有針對性、標準化和劑量依賴性的長期研究。
筆記和引用
3 Shigenaga M. K. Hagen T. M.和Ames B. N., 1994,氧化損傷與線粒體衰變,PNAS91(23), 10771 - 8。
4賽斯,H., 1997,氧化應激:氧化劑和抗氧化劑,Exp雜誌82(2), 291 - 5。
5王曉燕,王曉燕,王曉燕,1995,黃嘌呤-黃嘌呤氧化酶對黃萎病的影響2 +人類內皮細胞的儲存和蛋白質合成,中國保監會Res76(3), 388 - 95。
6 Dreher, D.和Junod, A. F, 1995,超氧化物、過氧化氫和羥基自由基對人內皮細胞內鈣的影響,J細胞雜誌162(1), 147 - 53年。
7 Harris, E. D, 1992,抗氧化酶的調節,美國實驗生物學學會聯合會J6(9), 2675 - 83。
8 Harris, E. D, 1992,銅作為銅、鋅超氧化物歧化酶的輔助因子和調節因子,J減輕122(3), 636 - 40。
9 Droge, W., 2002,自由基在細胞功能生理控製中的作用,雜誌牧師82(1), 47 - 95。
10 Simkó, M, Fiedeler, U., Gazsó, A.和Nentwich, M., 2008, Einfluss von Nanopartikeln auf zelluläre Funktionen。NanoTrust-Dossiers Nr. 007en, hrsg.;v. Institut für Technikfolgen-Abschätzung,維也納。
11 Simkó, M., 2007,細胞類型特定的氧化還原狀態負責不同的電磁場效應,咕咕叫地中海化學14(10), 1141 - 52。
12王曉明,陳曉明,陳曉明,陳曉明,陳曉明,2006,sind合成物是否為納米微粒?, NanoTrust-Dossiers Nr. 002, hrsg。v. Institut für Technikfolgen-Abschätzung,維也納。
13 Oberdorster, G., Oberdorster, E.和Oberdorster, J., 2005,納米毒理學:從超細顆粒研究演變而來的新興學科,環境衛生教諭113(7), 823 - 39。
14 Brown, D. M, Donaldson, K., Borm, P. J, Schins, R. P., Dehnhardt, M., Gilmour, P., Jimenez, L. A.和Stone, V., 2004,鈣和ros介導的超細顆粒暴露巨噬細胞轉錄因子激活和tnf - α細胞因子基因表達,m J Physiol肺細胞286 (2), L344-53。
15 Risom, L., Lundby, C., Thomsen, J. J., Mikkelsen, L., Loft, S., Friis, G.和Moller, P., 2007,急性缺氧和再氧誘導的人單核血細胞DNA氧化,Mutat Res625(1 - 2), 125 - 33所示。
16 Sioutas, C., Delfino, R. J.和Singh, M., 2005,大氣超細顆粒(UFPs)暴露評估及其在流行病學研究中的意義,環境衛生教諭113(8), 947 - 55。
17 Stone, V., Tuinman, M., Vamvakopoulos, J. E., Shaw, J., Brown, D., Petterson, S., Faux, S. P., Borm, P., MacNee, W., Michaelangeli, F.和Donaldson, K., 2000,暴露於超細炭黑時單核細胞細胞係鈣流入增加,歐元和J15(2), 297 - 303。
18 Poland, c.a, Duffin, R., Kinloch, I., Maynard, A.,Wallace, W. A., Seaton, Stone, V., Brown, S., Macnee, W.和Donaldson, K., 2008,引入小鼠腹腔的碳納米管在初步研究中顯示出石棉樣的致病性。Nat Nanotechnol3(7), 423 - 8。
來源:納米信托,奧地利科學院。NanoTrust檔案不定期發布,包含技術評估研究所在其研究項目NanoTrust框架下的研究成果。